
Urban Bird Diversity as an Indicator of Human Social Diversity and Economic Inequality in Vancouver, British Columbia
by Stephanie J. Melles
Department of Zoology, University of Toronto, Toronto, Ontario, Canada, M5S 3G5
Abstract
The unequal distribution of wealth in cities contributes to other forms of spatial, social, and biological inequities in complex, interacting, and self-reinforcing ways. Recent work on urban birds has often focused on community-level correlation studies of short duration in which many points along an urban gradient are surveyed for birds, and the data are related to various ecological variables measured at multiple scales. Spatial variation in urban bird communities may also reflect socioeconomic variables and cultural differences among the human population. The purpose of this paper was to examine whether socioeconomic factors (such as mean family income and ethnic diversity) also relate to the diversity and abundance of birds in Vancouver, British Columbia. I used redundancy analysis to characterize the socioeconomic gradient in a citywide study of the bird community in 44 census-defined neighborhoods. Mean family income, census tract area, and ethnicity were some of the dominant variables that correlated with most of the variation in the bird community. I found no direct relationship between neighborhood age and bird diversity and abundance. Results demonstrate that wealthier neighborhoods have more native species of birds and that these native species increase in abundance as the socioeconomic status of the neighborhood improves. With two-thirds of the world's population expected to live in cities by 2030, more and more people will grow up surrounded by a depauperate community of birds, and this could adversely affect the way people perceive, appreciate, and understand nature. Ultimately, as city birdlife diminishes and urban dwellers become dissociated from the natural diversity it represents, popular support for preserving and restoring such diversity may wane, allowing ecological conditions to further erode.
Key words: biodiversity; gradient analysis; mean family income; socioeconomic variables; spatial segregation; urban ecology
Introduction
"The fostering of wildife areas in cities is too complex an operation to be left in the hands of ecologists." — O.L. Gilbert
Of the three leading causes of species endangerment (urbanization, agriculture, and interactions with nonnative species), urbanization ranks highest (Czech, Krausman & Devers, 2000; Czech & Krausman, 1997). The process of urbanization endangers species by directly replacing native habitats with development on the urban-rural fringe, and because resources in the surrounding areas are depleted to support urban economies (Czech et al., 2000). Moreover, urban areas are expected to grow substantially in coming years: By 2030, the percentage of the world's population living in urban areas is projected to increase from the current 49% to approximately 61% (United Nations, 2004). Already in the United States, up to 80% of the population lives in suburban and urban areas (Blair, 2004; Grimm, Grove, Pickett & Redman, 2001). Urban sprawl may occur even faster in developing nations currently rich in biodiversity due to improving socioeconomic conditions (Liu, Daily, Ehrlich & Luck, 2003). As a result of continuous urban encroachment on natural habitats, the majority of the earth's human population will likely be living in a state of "biological poverty" by the year 2030 (Turner, Nakamura & Dinetti, 2004).
Biological poverty occurs when urban citizens experience below-average levels of native species diversity on a daily basis (Clergeau, Mennechez, Sauvage & Lemoine, 2001; Turner et al., 2004). Research on birds in cities worldwide has been steadily accumulating, particularly over the last few decades, and results indicate that as development intensifies, bird communities become increasingly homogenized (McKinney & Lockwood, 1999). Species richness and evenness also declines in complex ways, while total bird densities increase (Campbell & Dagg, 1976; Donnelly & Marzluff, 2004; Edgar & Kershaw, 1994; Emlen, 1974; Lancaster & Rees, 1979; Turner et al., 2004). Birds are often used as a biological model because they are good ecological indicators and they are easily observable (Clergeau et al., 2001). Moreover, trends that hold for birds may hold for other species of wildlife as well.
In the past, ecologists paid little attention to urban ecosystems and focused mainly on pristine ones (Blair, 2004; Collins, Kinzig, Grimm & Fagan, 2000; Jules, 1997; Marzluff, Bowman & Donnelly, 2001; Vandermeer, 1997). But ecological studies in urban areas now seem to be on the rise (Grimm et al., 2001). Much recent work by avian ecologists has focused on community-level correlation studies of short duration, in which many points along an urban gradient are surveyed for birds (Table 1). These kinds of studies are informative and cost effective as a first step, but unless we are able to identify all the processes that generate bird-community patterns (Table 1), our efforts to influence policy and planning will be largely ineffectual (Hostetler, 2001). A lot of interesting and challenging work remains to be done.
Experimental studies that focus on the underlying biological processes that drive ecological patterns are costly and intensive—and thus often limited to examinations at relatively small spatial scales, with low sample sizes. Moreover, they tend to focus on population-level effects rather than community interactions. Hence, some combination of mensurative and experimental work, done at a variety of spatial scales, is necessary (Table 1). Furthermore, to have a greater influence on urban planning and policy, human socioeconomic factors must be integrated into our investigations of diversity in urban areas; indeed, several researchers have suggested ways to do this (Dow, 2000; Grimm et al., 2001; Grove & Burch, 1997; Hope et al., 2003; Luck & Wu, 2002; Martin et al., 2004; Marzluff et al., 2001; McIntyre et al., 2000; Pickett et al., 1997; Pedlowski et al., 2002; Turner et al., 2004; see also Table 1).
Following research by Melles, Glenn, and Martin (2003), I examined here whether socioeconomic factors related to the bird community in Vancouver, British Columbia. Specifically, I tested whether human socioeconomic variables, such as wealth and human density, related to a gradient of avian diversity and abundance in Vancouver. By examining relationships with the dominant period of house construction in a neighborhood, I also tested whether "time since disturbance" created a habitat gradient that was related to the community of birds. Finally, I examined whether there was a clear pattern of ethnic spatial segregation that corresponded to patterns evident in the bird community.
Melles et al. (2003) investigated how the bird community changed along a gradient of increasing urban development. Bird abundance and diversity were investigated in relation to ecological attributes (for example, tree cover and impervious surface cover, composition, and number of tree species) measured at multiple scales (from 50 meters to 1 kilometer around point-count stations). But ecological factors alone cannot fully explain the patterns we see in urban areas: Humans are profound and industrious agents of change, and their behaviors influence ecological processes in cities. Other studies have found significant relationships between economic status and the type and distribution of plant species planted (Hope et al., 2003; Martin et al., 2004), as well as the type and spatial distribution of street trees (Pedlowski et al., 2002). Given that birds respond to the spatial heterogeneity and distribution of vegetation (Donnelly & Marzluff, 2004), the economic status of an urban neighborhood should have some relation to its bird community.
Methods
Study Area
My research was conducted in the municipalities of Vancouver, Burnaby, and Coquitlam, hereafter referred to as Greater Vancouver, in British Columbia, Canada (49°18' N, 123°12' W; Figure 1). These municipalities are located within the Vancouver Census Metropolitan Area (VCMA), a land area 2,412 square kilometers in size, which includes all surrounding municipal areas (such as Burnaby, Coquitlam, Surrey, Richmond, and North Vancouver). According to population and dwelling counts conducted by Statistics Canada (1996), the VCMA has a combined population of over 1.83 million people, and the average density of individuals in the area is 7.6 people per hectare. The maximum population density in the 44 census neighborhoods examined was 262.8 individuals per hectare and the minimum density observed was 4.4 individuals per hectare. The VCMA was the fastest-growing municipal area in Canada between 1991 and 1996, showing an increase of 14.3% in the overall population, and this growth is expected to continue. The VCMA is a largely urban and suburban area interspersed with several large parks (Figure 1).
Click image to enlarge
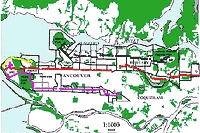
Figure 1: Study area, parks (green areas), census tracts (black outlines), and transect locations (four altogether, represented by purple, yellow, red, or black dots) for breeding bird and socioeconomic data in the Greater Vancouver area, British Columbia, Canada.
The term "urban," though quite common in everyday usage, can be somewhat subjective. Various organizations and individual researchers have defined it using such criteria as human population density, or the ratio of built (impervious surface cover) to unbuilt area (see, for example, Marzluff, Bowman & Donnelly, 2001). McIntyre, Knowles-Yánez, and Hope (2000) argue that there is a need for a working definition of urban that integrates ecological and social definitions, including baseline information about physical geography, demography, and socio-economic and cultural factors.
Here, I use Statistics Canada's census dictionary definition of the term "urban" (incidentally, I use "city" synonymously with "urban"). In this formulation, an urban area is defined as an area with a minimum population concentration of 1,000 individuals and a population density of at least 400 per square kilometer (Statistics Canada, 1996). "Urban cores" are areas with a population size greater than 100,000 individuals (i.e., Vancouver and the surrounding suburban municipalities of Burnaby and Coquitlam; see Figure 1), and "suburban areas" are defined as politically separate municipal areas located on the periphery of urban cores.
The physical geography of Greater Vancouver includes several large parks (> 3 km²) transected by road and trail systems that are used by many urban residents. The parks' relatively small impervious surface cover (between 2% and 35%) primarily results from the road network.
Neighborhoods surrounding these parks have a range of population densities, mean family incomes (see Figure 2), and varying levels of impervious surface cover (from 12% at the park edge to 70% in surrounding residential areas). Impervious surface cover in suburban areas varies between 34% and 70%, with a mean of 53%.
Click image to enlarge
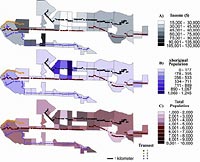
Figure 2a–c. : Census-tract neighborhoods in Greater Vancouver depicting a) trends in mean family income levels (20% of sample data), b) aboriginal population densities in 1996, and c) total population densities in 1996.
Vancouver is a young city—in the 1880s, it was a small settlement of sawmills, houses, and forest clearings surrounded by continuously forested land (Oke, North & Slaymaker, 1992). The city has expanded to its current size over the past 125 years and is expected to continue to grow. The ocean and mountains have tended to constrain or direct Vancouver-Burnaby urban development. A consequence of development has been the complete removal of forest and ground cover. Compensating to a small degree, street trees have been planted along many city streets. Initially, the west end of Vancouver was planted with native tree species such as Douglas fir (Pseudotsuga menziesii) and western hemlock (Tsuga heterophylla) (Oke et al., 1992). However, because they grew too fast and their root systems buckled the sidewalks, these species were eventually deemed unsuitable for urban settings. Thus, as the city grew eastward and southward, the trees planted were largely nonnative species, over a third of them nonnative cherries or plums (Prunus species). The result was an uneven distribution and composition of trees: In the newer suburbs on the west side of the city, the trees were often larger and more likely to be native compared to those on the east side, which had more deciduous, nonnative species (Oke et al.). This distribution is still evident today, although more diverse street tree plantings have begun to replace planted monocultures.
Some of the original vegetation of the area is retained in Greater Vancouver's large park system, and this resembles the dense coniferous forest of the coastal western hemlock (CWH) zone, with its shrub-dominated understory. The climax vegetation of this zone is generally dominated by a canopy of western red cedar (Thuja plicata) and western hemlock, with Douglas fir in drier areas and smaller numbers of Sitka spruce (Picea sitchensis), yellow cedar (Chamaecyparis nootkatensis), and lodgepole pine (Pinus contorta). Many species of nonnative vegetation, such as English holly (Ilex aquifolium) and Himalayan blackberry (Rubus discolor) are now also common in many Greater Vancouver parks and urban areas.
Bird Surveys
I collected relative abundance data for individual bird species at 285 point-count locations along four roadside transects in Greater Vancouver (see Figure 1; Melles, 2001). Point-count locations (with an interstation distance of 250 meters) were sampled once each year during the breeding season, 24 June–13 July 1997 and 1 May–1 June 1998, to maximize the number of sites that could be surveyed over the landscape. Birds flying over the point-count stations were not recorded because they were considered unlikely to be breeding in the area. The fixed-radius methodology (50 meters) was followed, and birds were recorded for a period of five minutes (see DeGraaf, Geis & Healy, 1991; Ralph, Geupel, Pyle, Martin & DeSante, 1993). All bird surveys were conducted on clear days during the first four hours following sunrise, to coincide with peak singing activity. I combined the bird-community data for 1997 and 1998 by selecting the maximum abundance for the two years at each point-count station. These abundance data were then averaged over all point-count stations within a given neighborhood census-tract area. Although the maximum value may be an optimistic estimate, it is likely to be a more accurate estimate of abundance at a particular site than the mean of one survey in each of two years (see Vander Haegen, Dobler & Pierce, 2000).
Because point-count data were only collected once during the breeding season of each year, 1997 and 1998, and the timing of data collection coincided with spring migration in 1998, this may have biased the abundance data in 1998. Some of the birds counted may have been migrants passing through the area. The focus in this study was on resident species of birds; thus, only species found in more than 10% of the census-tract neighborhoods and known to breed in the area were included in the analyses (n = 23 species). Though it is still possible that some of the individual birds recorded in 1998 were migrants, there were no significant year effects found between the community of birds recorded in 1997 and 1998 (data not shown, Melles, 2001).
Socioeconomic Data
According to Statistics Canada census definitions, a census family refers to a married or common-law couple (with or without children) or a lone parent of any marital status (Statistics Canada, 1996). I selected mean family income and the number of people holding a university degree (bachelor's or higher) from the 1996 Statistics Canada census as a measure of socioeconomic status. Of course, a number of economic variables could have been selected, but many were highly correlated, and these two variables should capture a large amount of the variability in both the income of an average residence and the education of an average person living in such a residence. I estimated neighborhood population density, spatial segregation among social classes, and racial composition using the census-tract data. All socioeconomic census data represented 20% of the total census population, whereas population estimates were absolute numbers of people.
I used the number of houses constructed during different time periods to estimate time since neighborhood development. This type of variable has been used before in a related study (Martin et al., 2004) to estimate the "time since disturbance," and it assumes that prior to the development, the area would have retained some of the original native vegetation. Logging operations may have cleared some of the land prior to neighborhood development, but the disturbance caused by construction of houses and impervious roadways removes land from natural regenerative processes. Thus, it is reasonable to estimate time since disturbance by the number of houses constructed in a census neighborhood during a given period of time.
Data Analysis
To characterize the relationships between socioeconomic status, time since disturbance, spatial segregation among ethnic backgrounds, and the abundance of different bird species one might expect to see in a given neighborhood, I used redundancy analysis (RDA) (ter Braak & Šmilauer, 1998; Legendre & Legendre, 1998). RDA is akin to direct gradient or regression analysis done in multivariate species space (ter Braak & Šmilauer). RDA relates abundance data from a species matrix to a matrix of environmental data using multiple linear regression techniques, and it assumes that species have linear responses to ecological gradients. That is, the abundance of a species is expected to increase linearly along a gradient. Linear responses may arise when species distributions extend beyond the extremities of the gradient sampled (Austin, 2002).
I selected redundancy analysis as opposed to canonical correspondence analysis (CCA) because a preliminary examination of the data showed that the length of the species gradient was short with respect to the socioeconomic variables (gradient length = 1.91), and RDA is recommended when gradients are short (i.e., < 3 SD; ter Braak & Šmilauer, 1998). The species matrix consisted of the average abundance per census-tract area of 23 species of birds (Table 2). Although 48 species were detected in the study area, only birds present in more than 10% of the 44 census neighborhoods were included in the analysis in order to exclude potential migrants that were not breeding in the area (as noted above). The final species matrix was related to a linear combination of 12 socioeconomic variables (Table 3). The significance of the RDA ordination of species and human socioeconomic relationships was investigated by performing a randomization test on the projected relationships (ter Braak & Šmilauer); 199 random permutations were performed on the significance of the ordination axes. Randomizations were spatially restricted by the linear transects in order to ensure that shuffling was not entirely random across the study area. Data were not sampled randomly across the study area, and hence permutations were restricted within the line transects (CANOCO 4; ter Braak & Šmilauer). Redundancy analysis allows one to infer which variables best explain the variation in species distributions because the most important variables load highest on the first axis.
Variance Partitioning
Census tracts in Greater Vancouver did not cover equal amounts of area (Figure 1), and this could result in some census neighborhoods having more species simply because the area sampled was larger. Variance partitioning is a technique used to examine the relative contribution of different factors while controlling for covariables that may have overlapping effects (Bocard, Legendre & Drapeau, 1992). I was interested in partitioning out the amount of variation in the species data that could be attributed to the area of a census neighborhood. In addition, I wanted to determine the relative contribution of another potentially confounding factor—the spatial structure of the species data (Bocard et al., 1992). As suggested by Bocard and colleagues, spatial structure was modeled using trend surface analysis. I used the spatial coordinates of the census-tract centroids, defined using the universal transverse mercator, North American datum 83 projection, as covariables in partial RDA analyses. Only the X and Y coordinates were used because all higher-order terms (for example, X², XY) were highly correlated with these two. I used variance partitioning to partition out the amount of variation in the species data that could be attributed to socioeconomic variables, spatial variables, and area (Bocard et al., 1992; Cushman & McGarigal, 2002).
Results
A map of Greater Vancouver depicting the study area (Figure 1) shows neighborhoods (delineated as census tracts outlined in black) in relation to their proximity to large urban parks. Figure 2 (a–c) shows that census-tract neighborhoods with the lowest mean family incomes (2a) are also the areas with the highest proportion of people of aboriginal ethnicity (i.e., North American Indian, Métis, or Inuit and/or those who reported being a Treaty Indian or a Registered Indian as defined by the Indian Act of Canada) (2b). These neighborhoods also have the highest population densities in Greater Vancouver (2c), have the fewest small parks, and are the farthest away from large urban parks (Figures 1 and 2).
Click image to enlarge
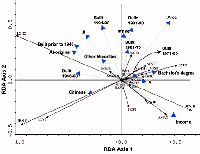
Figure 3: Redundancy analysis, ordination bi-plot examining the strength of association between human social variables and 23 bird species in an urban bird community, Greater Vancouver, BC. (See Table 2 for bird name abbreviations.) Species and socioeconomic variables are represented by arrows that indicate the direction in which the variables are increasing.
Mean family income had the strongest positive correlation with RDA axis I (Figure 3; Table 3, interset correlation with RDA I = 0.47), indicating that this variable has a strong influence on the separation of the bird species data along a socioeconomic gradient; the majority of native bird species were positively related to increasing socioeconomic status. Mean family income was followed closely by census-tract area (Figure 3; Table 3, interset correlation with RDA I = 0.43). I expected that area would be a strong explanatory variable given that the species-area relationship is one of the most general patterns found in ecology. In short, as the logarithm of the area sampled increases, more and more species are detected (Pileou, 1966).
The positive relationship between the number of people holding university bachelor's degrees (or higher) and native avian species diversity (Figure 3; Table 3, interset correlation with RDA I = 0.30) likely reflects the location of Vancouver's university campuses. Both the University of British Columbia and Simon Fraser University are surrounded by park space and university endowment lands. So the diversity of birds in these areas reflects park habitat rather than people's personal preferences or advanced learning shaping the local habitat of their neighborhood.
Two other interesting findings can be ascertained from the RDA ordination (Figure 3). First, there appears to be no discernable relationship between time since disturbance and the distribution of bird species, contrary to expectations. Although several of the disturbance variables were significantly correlated with the RDA axes (Table 3), there is no apparent trend through time—the community of birds in neighborhoods built primarily prior to 1946 is similar to the community of birds near newer houses built between 1990 and 1996. But older neighborhoods have more nonnative species of birds (Figure 3). The second noteworthy finding is that persons with aboriginal ethnicity reside predominately in areas with fewer bird species, and the bird species that do occur in these areas are generally nonnative. This finding further describes the spatial segregation indicated in Figure 1.
Click image to enlarge
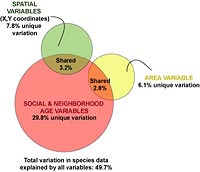
Figure 4: Variance decomposition based on partial redundancy analyses of the bird community in Greater Vancouver, BC. Pink represents variance explained by social and neighborhood-age variables, after removing the effects of covariates area and space (XY coordinates). Yellow represents variance explained by census-tract area alone, after removing the effects of the space covariate and social + neighborhood-age variables. Green represents variance explained by space alone, after removing the effects of the area covariate and social + neighborhood-age variables. Amounts of shared variance were calculated by running a further series of partial redundancy analyses and calculating the joint or shared effects of 1) social + neighborhood-age variables, constrained by area, 2) social + neighborhood-age variables constrained by space, and 3) area, constrained by space.
Variance partitioning was used to determine how much of the variation in the distribution of bird species can be attributed to either the area of the census tract surveyed or the tendency for bird species distributions to have some level of spatial structuring. Figure 4 shows that the majority of the variance in the urban bird community of Greater Vancouver can be explained by socioeconomic and time since disturbance variables (neighborhood age was grouped with the social variables). Almost 50% of the variation in bird abundance data can be attributed to all three factors (social, area, and space) combined. However, social variables account for 29.8% of that variation; census area accounts for 6.1%; and spatial structuring in the bird community accounts for almost 7.8%. Only 6.1% of the variation in the avian community is shared between socioeconomic variables and either spatial variables or area. Census area (hectares) and the two spatial variables did not share any amount of variation. Although social variables and time since disturbance were able to explain more variance, we should be cautious about interpreting this to mean that these variables are better or the more important factors in the system. That's because there were four times more social and disturbance variables than area and spatial variables combined (see Table 3).
Discussion
According to Turner, Nakamura, and Dinetti (2004), most of the world's human population lives in biological poverty. In one study examining the relationship between human population density and species diversity at global scales, they examined data from five cities around the world and found an inverse relationship between the numbers of humans and the diversity of birds in neighborhoods (scaled at one square kilometer). My work in Greater Vancouver substantiates their findings from Berlin, Germany; Washington, D.C., USA; Florence, Italy; Chiba, Japan; and Tucson, Arizona, USA. (Only 6 of 23 species of birds in Greater Vancouver were related to increasing numbers of humans; see left-hand side of Figure 3). But the bird-human relationship is not necessarily straightforward in Greater Vancouver: Complex socioeconomic and cultural factors are also correlated with the diversity of birds. Neighborhoods of higher socioeconomic status tended to have more native species of birds than ones of lower socioeconomic status (i.e., those predominantly composed of aboriginal peoples). Furthermore, Turner, Nakamura, and Dinetti (2004) argue that human ability to assess the overall ecological health of an area diminishes as new generations are exposed to poor ecological conditions. If this is indeed the case, then certain city neighborhoods with persistent low socioeconomic status are in danger of becoming self-perpetuating and self-segregating areas of low biodiversity.
It is perhaps not surprising that socioeconomic status was strongly correlated with the community of birds in Greater Vancouver (Figure 3). If socioeconomic status limits a family's ability to purchase a house in the neighborhood of a large park, then this indeed follows from that expectation. Melles, Glenn, and Martin (2003) showed that park area and coniferous and deciduous tree cover (within one kilometer of avian point-count stations) were significantly related to the likelihood of observing most native species of birds. Even the likelihood of finding more common species like the American robin (Turdus migratorius) significantly increased with the amount of park area in the vicinity. Greater Vancouver is relatively unique (and fortunate) in having large remnants of historic vegetation; however, the unequal spatial distribution of these remnants constrains access to these large parks. Land values in urban areas are often heavily influenced by the proximity of parks and other green space. Affordable urban housing is often limited to abandoned commercial and industrialized areas of the city, where there is more impervious surface and less green space.
I did not find a relationship between time since disturbance and the bird community, though I expected that older neighborhoods would have more well-advanced vegetation (larger trees and shrubs) and thus higher bird species richness and abundance. Upon closer inspection of the data, it became clear that the amount and type of vegetation in older areas could not be generalized. One of the oldest neighborhoods in Vancouver is located close to the downtown core, just to the east of Stanley Park (Figure 1, labeled "Historic A"). Because this area is so close to the urban center, the density of buildings is high, and much of the vegetation has been removed, leaving little habitat for birds. Another historic neighborhood in Vancouver is located in the west end (Figure 1, labeled "Historic B"). Here, many large mature street trees—native species such as Douglas fir and western hemlock—remain standing today and provided habitat for a variety of bird species. Several neighborhoods to the east of the downtown core, developed primarily between 1946 and 1960 (data not mapped), were initially planted with many nonnative cherry and plum street trees. These ornamental fruit trees have short life spans and require replacement, and as such, these plantings have not been conducive to maintaining a diverse bird community. Therefore, contrary to my expectations, older neighborhoods did not necessarily have a more well-advanced vegetation community in Greater Vancouver.
Landscape-scale spatial heterogeneity in urban areas is established and maintained in a "top-down" way by formal institutions like city planning departments (through zoning bylaws), public works, and courts (Grimm et al., 2001). In Vancouver, there are historic reasons for the spatial distribution of parks (for example, federal endowments of land to the universities and the historic transfer of lands previously held in federal government reserves, such as for Stanley Park; see Figure 1). However, less formal "bottom-up" actions such as tree plantings, community gardens, and park maintenance by community groups, families, and associations can have a profound influence at the local level and also contribute to large-scale spatial heterogeneity. Residential and local community gardens planted with large berry-producing shrubs (e.g., salmonberry, blackberry, and elderberry) and conifer trees, for instance, had a higher likelihood of being occupied by many species of birds in Greater Vancouver, including bushtits (Psaltriparus minimus) and spotted towhees (Pipilo maculatus) (Melles et al., 2003). It must be noted, however, that impoverished areas tend to have lower levels of residential involvement in neighborhood tree planting and community efforts, and this could reinforce social and spatial segregation (Pedlowski et al., 2002).
Social scientists have long examined how human perception, choice, and action drive the political, economic, and cultural decisions that lead to—and respond to—changes in urban areas (Jacobs, 1961; Grimm et al., 2001). Why is there such extreme evidence of ethnic segregation in Greater Vancouver, particularly with respect to the aboriginal population? And what does the unequal distribution of environmental amenities mean in terms of social justice? These two questions reflect some of the most complex issues affecting contemporary urban life (Pedlowski et al., 2002). Jacobs (1961) argues that some of the factors that foster healthy and ethnically diverse neighborhoods in urban areas are community involvement, low rent-to-income levels, cultural and spatial heterogeneity, and active social interactions at the street level. In Greater Vancouver, there are a variety of other historic and social factors that also influence the extreme patterns we see. In order to turn these patterns around, we need to see far greater social, economic, scientific, and community involvement in neighborhoods that are often ignored or avoided.
Understanding why certain neighborhoods are ignored by city planners and avoided by urban residents is the first step to changing these patterns. Ethnic and economic segregation exists in many cities worldwide, but in Canada it is especially evident in Vancouver. It is a complex and systemic problem involving many factors such as intravenous drug and alcohol abuse, poverty, homelessness, and neglect. Many of these problems are most rampant among aboriginal people (Riley, 1998). The following statistic demonstrates just how bad the situation can get: Vancouver's east side set the world record in 1998 for the largest increase in the number of HIV cases among intravenous drug users (Riley, 1998; Nolin, 1999).
Often there is a social stigma associated with poverty and with people who are addicted to drugs and alcohol, as though they have brought their circumstances and illnesses upon themselves (Nolin) and as though their position bears no reflection on society as a whole. This sort of social stigma perpetuates ethnic spatial segregation. Many people refuse to admit that drug consumption is a health problem (Nolin), even as the costs for Canada (in 1992) were estimated at more than $18 billion (Riley), including the combined costs of drug enforcement, loss of productivity due to morbidity, premature deaths, and direct health costs. Recent initiatives under the city of Vancouver's draft plan "for the prevention of harm from psychoactive substance abuse" (2005) hold considerable promise for turning these patterns around. This plan aims to prevent harm by approaching the problem from the perspective of population health, adequate housing and employment, access to health care, and individual and community-based approaches.
Interestingly, approaches designed to prevent harm from substance abuse are the same sort of top-down and bottom-up approaches that could prevent the further deterioration of urban ecological health. For example, creating healthy school environments (including those of schoolyards) and supporting parents to help engage youth in community and social initiatives like urban renewal should have positive ecological and health outcomes. Research has shown that concern for natural features can be fostered by direct interaction with such features (Turner et al., 2004; Bixler, Floyd & Hammitt, 2002; Schultz et al., 2005). Ultimately, social health may be inextricably linked to ecological health.
People living in the poorest neighborhoods, at the extreme end of the socioeconomic gradient, have lower levels of residential involvement in neighborhood tree planting and community green-up efforts than better-off citizens, probably because they are more concerned with the immediate issues of day-to-day survival. Although biological poverty in these neighborhoods does have implications for environmental justice, it is unlikely, in the near term, that fewer species of birds in these areas will lead to reduced support for the protection of biological diversity in Vancouver: The numbers of people in this very vulnerable segment of society are relatively small here (City of Vancouver, 2005).
However, as more and more people in middle- to low-income neighborhoods grow up surrounded by a depauperate bird community, the ability to assess ecological health may indeed diminish. Urban citizens perceive and appreciate the "nature" they know, so as new generations are exposed to poor ecological conditions, the dissociation from biological diversity could lead to lower popular support for natural diversity at regional and even national levels. With increasing population growth in urban neighborhoods, it is becoming more important to plan for the kinds of wildlife values we wish to uphold at municipal, provincial or state, and national levels. Ad hoc urban planning, with no attention paid to the importance of vegetation and native wildlife for overall community health, will lead to environmental erosion. Though several community green-up initiatives do stem from bottom-up groups, these are unlikely to be able to deal with the magnitude of the problem. It is reasonable to conclude that minimum standards and bylaws need to be set for such amenities as native tree and shrub plantings and removals, as well as the maintenance of drought-resistant residential garden plants.
In terms of scientific involvement, there are a number of logistical and social obstacles associated with doing research in urban habitats, and this is perhaps why experimental studies are so scarce (Table 1). First, gaining access to numerous private properties can be a daunting task for scientific researchers, especially younger ones, who are perhaps not comfortable communicating and justifying their research to the public. Second, there are issues of privacy and safety: Some people might not want a researcher peering with binoculars into their backyard. Third, there is inadequate financial support and political backing for the study of urban areas, possibly because of the attitude that there are more deserving areas for our conservation and research dollars. Fourth, urban areas are extremely altered, complex systems that provide coincidental habitat for wildlife in an environment designed primarily to provide a variety of socioeconomic services that satisfy human needs, preferences, and desires. Wildlife species in urban areas are viewed as incidental (Clergeau et al., 2001) and perhaps even nonessential inhabitants. Birds may indeed be a source of pleasure for people in urban, suburban, and rural areas alike (Clergeau et al., 2001), but in urban areas they often go unnoticed.
Acknowledgments
Thanks to Marie-Josée Fortin for her insightful comments and encouragement to pursue this research. Dr. Fortin provided valuable support and editorial guidance on earlier drafts of the manuscript. Thanks also to two anonymous reviewers and the journal editors, whose helpful suggestions improved the manuscript considerably. This project was funded with support from the Asa-Yohal Graduate Fellowship in Forestry and the Bert Hoffmeister Scholarship in Forest Wildlife. The Centre for Applied Conservation Biology, the Geography Department at the University of British Columbia, the Zoology Department at the University of Toronto, and the Canadian Wildlife Service also provided many resources and support. I would also like to gratefully acknowledge Brian Klinkenberg for motivating discussions of some of the ideas presented in this paper. Thanks also to the citizens of Greater Vancouver who provided valuable information in their Statistics Canada census-response forms and allowed me to observe and record the birds and vegetation in their yards.