
Avian Response to Restoration of Urban Tidal Marshes in the Hackensack Meadowlands, New Jersey
by Alison Seigel,¹ Colleen Hatfield,² and Jean Marie Hartman³
¹Rutgers University, Graduate Program in Ecology and Evolution, 1 College Farm Road, New Brunswick, NJ 08901
²California State University, Department of Biological Sciences, 400 West First Street, Chico, CA 95929 (address at time of research: Rutgers University, Department of Ecology, Evolution, and Natural Resources, 14 College Farm Road, New Brunswick, NJ 08901)
³ Rutgers University, Department of Landscape Architecture, 93 Lipman Drive, New Brunswick, NJ 08901
Abstract
Tidal marshes located in urbanized regions have experienced a long history of degradation. As a result, restorations have frequently been conducted to improve the habitat quality of these marshes. Few studies, however, have investigated the effect of restoration on avian community composition in urban tidal marshes. To this end, we conducted avian surveys for one year prior to restoration and three years after restoration at Harrier Meadow marsh, in the Hackensack Meadowlands, New Jersey. After restoration, avian species richness and abundance increased, while evenness decreased, mostly due to large flocks of sandpipers sporadically visiting the marsh during migration. Prior to restoration, generalists were by far the most abundant foraging guild, while they shared dominance with mudflat and open-water foragers after restoration. Avian surveys were also conducted for three years after restoration at Mill Creek marsh, also in the Meadowlands. Though the restoration goals were the same for Harrier Meadow and Mill Creek, the two marshes had distinct habitat compositions after restoration, and this allowed us to examine avian response to variation in habitat availability. In all three years of monitoring after restoration, Harrier had a greater avian density and higher species richness than Mill Creek; however, avian abundance at both marshes was dominated by the same three foraging guilds. Evenness did not differ across post-restoration years or between marshes. Avian abundance showed a decreasing trend during the three years of post-restoration monitoring; however, further monitoring will be necessary to determine the long-term trends in the avian community.
Key words: avian community structure, foraging guild, Hackensack Meadowlands, marsh restoration, Phragmites, restoration monitoring, tidal marsh, urban
Introduction
Estuarine tidal marshes serve as important foraging grounds and juvenile nurseries for a variety of fish species and crustaceans (Kneib, 1997) and provide important habitat for many migratory and resident bird species (Reinert & Mello, 1995; Burger, Niles & Clark, 1997; Melvin & Webb, 1998). Yet many coastal North American metropolitan areas, such as New York City, Boston, Seattle, and San Francisco, are located in, on, or near tidal marshes. As a consequence, urban tidal marshes have been altered for various types of development, and this has caused tidal restriction and habitat fragmentation and loss. Urban marshes have also been on the receiving end of a wide variety and high concentration of pollution from sources such as landfills and industry. Urban influences are believed to account for almost 60% of wetlands loss in the United States (Opheim, 1997).
In the northeastern U.S., the remaining urban tidal marshes are frequently dominated by Phragmites australis (common reed), a plant whose presence typically indicates an altered and degraded habitat (Winogrond & Kiviat, 1997; Chambers, McComb & Tappeiner, 1999). Few studies have focused on avian use of Phragmites-dominated marshes; however, it is generally thought that these degraded marshes provide limited resources for wetland birds (Roman, Niering & Warren, 1984; Benoit & Askins, 1999). The physical structure of dense, monospecific stands of Phragmites prevents shorebirds, waders, waterfowl, and other taxa from gaining access to the marsh surface for foraging.
The use of tidal-marsh restoration has increased in response to the continued degradation of wetlands and an enhanced understanding of the value of wetlands in urban areas (Zedler, 1996; Bergen, Alderson, Bergfors, Aquila & Matsil, 2000; Harbor Estuary Program [HEP], 2001). However, urban marshes have typically experienced a long history of perturbation and continue to be influenced by urban pressures. Under these circumstances, restoration potential is compromised, and evaluation of restorations based on comparison to conditions at reference sites may not be possible or appropriate. Instead, restorations can be evaluated using a same-site, "pre-restoration versus post-restoration" monitoring strategy. Use of this type of monitoring in tidal-marsh restoration has been rare, and thus it is an important focus for study.
Click image to enlarge
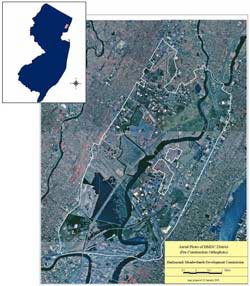
Figure 1a. Map of the New Jersey Hackensack Meadowlands District. Boundaries are indicated by white lines.
Much literature exists demonstrating the importance of tidal-marsh habitat to numerous bird species throughout the year (Chavez-Ramirez & Slack, 1995; Reinert & Mello, 1995). However, relatively few studies have focused on avian response to marsh restoration, and of these, only a small number have taken place in urban areas. This is despite the fact that in terms of restoration monitoring, birds may be particularly good indicators of habitat quality because they integrate multiple environmental influences in a habitat and respond quickly to changes in habitat (Neckles et al., 2002; U.S. Environmental Protection Agency [EPA], 2002).
The goal of this study was to evaluate avian response to tidal-marsh restoration in an urban landscape. The objectives were to 1) compare and contrast the pre-restoration and post-restoration avian community in a restored marsh; and 2) to compare temporal, post-restoration trends in the avian community at two restored tidal marshes in close proximity to one another.
Study Sites
The New Jersey Hackensack Meadowlands (the Meadowlands) is a mosaic of brackish and freshwater tidal wetlands, freshwater non-tidal wetlands, uplands, and developed areas that includes the largest remaining tidal-marsh complex (3,400 hectares) in the New York–New Jersey Harbor Estuary (Figure 1a). Seven miles west of New York City, the Meadowlands lies along the Atlantic Flyway and is surrounded by an urban matrix. Over 90% of estuarine marshes in the Meadowlands are dominated by Phragmites due to decades of land alteration that created conditions favoring invasion by this species (Sipple, 1972; Tiner, Swords & McClain, 2002). Two brackish marshes in the Meadowlands—Harrier Meadow and Mill Creek—were included in this study (Figure 1b).
Click image to enlarge
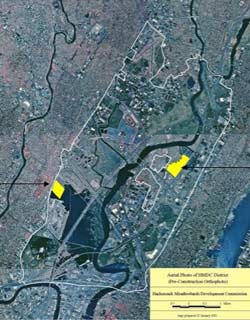
Figure 1b. Location of Harrier Meadow and Mill Creek marshes within the Meadowlands District.
Harrier Meadow is a 32.2-hectare tidal marsh surrounded by tidal mudflats on two sides and urban development and landfill on the remaining two sides (Figure 2). Prior to restoration, Harrier Meadow was dominated by Phragmites and Lythrum salicaria (purple loosestrife) and featured scattered, small pools of water and isolated patches of native high-salt-marsh vegetation, such as Spartina patens (saltmeadow cordgrass) and Distichlis spicata (saltgrass). Just under 22 hectares (71%) of Harrier Meadow were restored in 1998. The restoration design included the creation of three large, tidally influenced open-water areas surrounded by high-marsh and fringe-upland vegetation. The remaining area, which consisted of high-marsh vegetation and Phragmites, was not altered. Restoration of Harrier Meadow was intended to create and enhance a variety of habitats for wildlife and to bring about the recovery of wetland function (Hartman, 2002a).
Mill Creek is a 56.7-hectare tidal marsh bordered by the New Jersey Turnpike on the east and residential land use on the west (Figure 3). Prior to restoration, Mill Creek was dominated by Phragmites and contained very little open water. General restoration goals for Mill Creek Marsh were similar to those for Harrier Meadow (Hartman, 2002b). Thirty-eight hectares (67%) of Mill Creek were restored in 1999, creating a tidal channel, tidally influenced open-water areas, and mudflats with interspersed islands of upland vegetation.
Approximately the same percentages of the two marshes were restored; however, Harrier Meadow and Mill Creek had distinctly different habitat compositions after restoration (Table 1a). Almost a third of Harrier Meadow (mostly in the unrestored portion of the marsh) was still covered by Phragmites, while this species was a negligible habitat component of Mill Creek. Mudflat, open water, upland, high marsh (Harrier Meadow only), and low marsh (Mill Creek only) habitats were significant components of both marshes, but in different configurations. These differences represent one of the challenges in evaluating responses to restoration, in that there is rarely an opportunity to have true replicates in restoration design. In this study, comparison of two designs enabled us to evaluate the general avian response to marsh restoration because both marshes were located within the same habitat complex and drew avian species from the same regional species pool. We were also able to contrast how variation in habitat availability led to differences in avian response.
Click image to enlarge
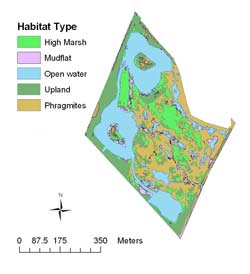
Figure 2. Distribution of six habitat types at Harrier Meadow Marsh after restoration. Though not illustrated here, the southeastern and southwestern sides of the marsh were adjacent to mudflats.
Methods
Habitat Analysis
Digital habitat maps of Harrier Meadow and Mill Creek were created with geographic information system (GIS) software, Environmental Systems Research, Inc.'s ArcInfo 9, using color infrared digital aerial photographs and detailed field observations. Area coverage of habitat types was calculated in ArcMap using the digital vegetation maps. Habitats at Harrier Meadow included high marsh, mudflat, open water, Phragmites, and scrub-shrub/upland (Table 1b). Habitats at Mill Creek included low marsh, mudflat, open water, Phragmites, and upland.
Avian Surveys
We conducted pre-restoration surveys of birds at Harrier Meadow in 1997. Mill Creek was not surveyed prior to its restoration due to the dense stands of Phragmites at the site, which severely limited the ability to conduct surveys. Post-restoration surveys were conducted at both marshes in 2001, 2002, and 2003. In all survey years, we surveyed each marsh five times during each of three seasons: spring migration (mid-March through mid-May), summer (early June through late July), and fall migration (early August through mid-October). Each marsh was therefore surveyed 15 times per survey year.
We conducted the avian surveys by scanning predetermined, fixed areas (stations) within each marsh and recording all individuals detected visually and audibly within a station during a five-minute period (see Seigel, 2006, for further explanation). Survey stations were delineated prior to restoration and did not change during monitoring. One avian survey consisted of a count at all stations present at the marsh. There were six survey stations at Harrier Meadow (four within the restored portion of the marsh and two in the unrestored portion) covering a total of 9.8 hectares. Included in the total survey area were 2.9 hectares located outside the marsh boundaries in the surrounding mudflat. Mill Creek contained five survey stations (all within the restored portion of the marsh) covering 14 hectares. Individual birds flying over the marsh were not recorded, with the exception of foraging raptors and aerial insectivores. Care was taken not to count the same individual twice. We surveyed the marshes between sunrise and 10 a.m. Surveys were not conducted in rain or heavy wind. Species such as bitterns and rails were underrepresented by the surveys because we did not use vocalization tapes for these species.
Click image to enlarge
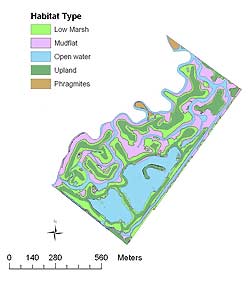
Figure 3. Distribution of four habitat types at Mill Creek Marsh after restoration.
Individual birds were identified to the species level. In three instances, however, individuals were identified to the genus level due to an inability to distinguish between species (American crow and fish crow, recorded as Corvus species) or difficulty in identifying individuals in large flocks (greater and lesser yellowlegs, recorded as Tringa species, and semipalmated and least sandpipers, recorded as Calidris species).
Analysis
We assessed avian response to restoration using two approaches. The first compared the pre- and post-restoration avian communities at Harrier Meadow. To keep sampling effort equal, we compared one year of pre-restoration data (1997) with one year of post-restoration data (2002), the middle year of the three-year post-restoration data set. Our second approach examined post-restoration (2001–2003) trends in the avian communities at both of the restored marshes. Metrics used to characterize the avian communities included avian abundance, species richness, diversity, evenness, and Sorensen's similarity.
We also studied foraging guilds to examine how the change in habitat structure at each marsh affected the avian communities. The avian community was categorized into six guilds: generalist, aerial, upland, Phragmites, open-water, and mudflat foragers (Table 2). Generalists included species that foraged in more than one type of habitat. Species were placed in a guild based on the predominant habitat used for foraging (based on Ehrlich, Dobkin & Wheye, 1988; personal observation). There was only one species (common snipe, Gallinago gallinago) that was observed foraging in high-marsh habitat, and therefore this habitat and associated species were not included.
We realize that our method of analysis does not contain true replication in avian response to restoration. However, with the exception of extremely well funded projects, replication is nearly impossible in this type of study. In any case, repeated, independent studies of numerous restorations are necessary for developing strong inferences regarding patterns and cause-effect relationships in avian response to restoration. While we understand the limitations of our approach for statistical analysis, the careful documentation this study represents is an essential first step toward developing a body of scientific research.
Click image to enlarge
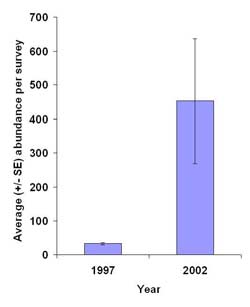
Figure 4. Average (± SE) avian abundance per survey at Harrier Meadow prior to restoration (1997) and after restoration (2002).
We took a very conservative approach to analysis by using descriptive statistics (mean ± standard error) to detect differences in avian community characteristics (abundance, species richness, diversity, and evenness) before and after restoration at Harrier Meadow, and between the restored Harrier Meadow and restored Mill Creek. Diversity of the avian communities was measured using the Shannon-Wiener diversity index (H'): H' = –Σ(pi) × (lnpi), where pi is the proportional abundance of species i, summed for all n species measured. Evenness is defined as the Shannon diversity divided by the maximum possible diversity (Krebs, 1989). Similarity of species composition of avian communities between two years was determined using Sorensen's similarity index (Cs): Cs = 2j / (a+b), where j is the number of species present in both years, a is the number of species in the first year, and b is the number of species in the second year (Magurran, 1988).
Because total survey areas at Harrier Meadow and Mill Creek were not equal, comparisons of avian community characteristics between the two marshes were standardized by factoring in the number of hectares surveyed. We divided average avian abundance by the number of hectares surveyed to obtain average density per survey. Species richness was also divided by the number of hectares surveyed and is presented as the number of species observed per hectare per survey. The species-richness-per-hectare measure should be considered with caution, however, as the relationship between species richness and area isn't necessarily linear.
Results
Pre-restoration Versus Post-restoration at Harrier Meadow
Through the addition of open water and other habitats during restoration, Shannon diversity of habitat types at Harrier Meadow increased from 0.965 in 1997 to 1.121 in 2002. A total of 43 bird species were observed in the marsh in 1997. In comparison, cumulative species richness in 2002 was 57, an increase of more than 30% after restoration. Similarity of species composition in the avian community before and after restoration was 0.62.
Average avian abundance per survey exhibited more than a tenfold increase, from 33 (± 3) prior to restoration to 453.80 (±184) after restoration (Figure 4). Red-winged blackbird (Agelaius phoeniceus) and sandpipers (Calidris species) were the most abundant species before and after restoration, respectively. Average avian species diversity per survey was essentially the same before and after restoration (Table 3). Changes to the elements of diversity—species richness and evenness—were more indicative of the influence of restoration. On a per-survey basis, pre-restoration species richness was approximately half that of post-restoration. Evenness per survey decreased by 24% after restoration.
Click image to enlarge
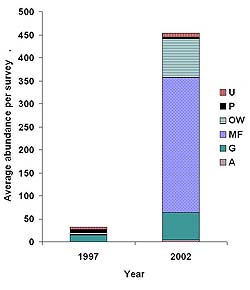
Figure 5. Average abundance per survey of six foraging guilds before (1997) and after (2002) restoration at Harrier Meadow. Guilds include upland foragers (U), Phragmites australis foragers (P), open-water foragers (OW), mudflat foragers (MF), generalists (G), and aerial foragers (A).
To determine if the large flocks that periodically visited the marsh masked a change in diversity after restoration, we removed sandpipers (Calidris species) from the analysis. Without the considerable reduction in evenness caused by the temporal variability of large sandpiper flocks, annual diversity was higher (2.11 ± 0.08) after restoration.
To examine how changes in habitat types after restoration influenced avian community structure, we compared the average abundance per survey of foraging guilds before and after restoration (see Table 4 for standard errors). Prior to restoration, all guilds were present in low abundance except for aerial foragers, which were absent (Figure 5). The dominant foraging guild was generalists. After restoration, average abundance per survey increased markedly in three of the six guilds: Mudflat foragers increased from 0.73 (± 0.05) to 294 (± 189), open-water from 5.5 (± 1.2) to 84.9 (± 12.4), and generalists from 14.9 (± 2.3) to 58.9 (± 18.7). The only guild to decrease in abundance after restoration was Phragmites foragers, which dropped from 5.6 (± 1.3) to 3.3 (± 0.8) individuals per survey.
Species richness within foraging guilds showed a pattern of post-restoration change generally similar to that seen in abundance (Table 5). Mudflat and open-water foragers showed the largest percent increase in species richness after restoration. Although three additional species were present after restoration in the upland foraging guild, seven species were absent, resulting in a net loss of species. Pre- and post-restoration within-guild species composition was similar for generalist, upland, and open-water foragers.
The marshes of the Meadowlands serve as important stopover sites during spring and fall migration and breeding habitat in the summer. Seasonal changes in avian community composition are masked when examining the effect of restoration only on the annual level. Therefore, we examined pre- and post-restoration seasonal patterns in the avian community. Prior to restoration, average avian species richness per survey did not vary a great deal by season, though there was a trend for species richness to decrease from spring to fall (Figure 6). In contrast, average species richness per survey increased considerably after restoration in the spring, summer, and fall. After restoration, fall species richness was lower than spring and summer because there were 22 species, mostly waterfowl and passerines, present in the marsh in spring and/or summer but not in fall. Before restoration, nine of 43 species were observed in all three seasons, and three species (0.07%) had a frequency ≥ 0.67. After restoration, 27 of 57 species were observed in all three seasons, while 12 species (21%) had a frequency ≥ 0.67 (Table 6).
Click image to enlarge
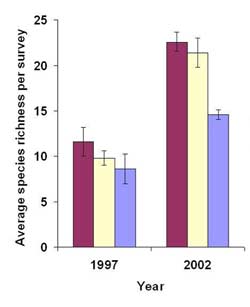
Figure 6. Average (± SE) avian species richness per survey before (1997) and after (2002) restoration at Harrier Meadow during fall and spring migration and summer breeding season.
Average diversity per survey was similar across seasons prior to restoration (Table 7). In contrast, post-restoration diversity decreased from spring to fall, and there was a tendency for greater variability in diversity in summer and fall. Pre- and post-restoration seasonal diversity were similar. Post-restoration spring had the highest diversity, primarily due to a doubling in species richness from pre-restoration accompanied by a relatively small decrease in evenness. Diversity in the fall decreased due to a relatively large decrease in evenness. The highest degree of seasonal variability in diversity accompanied the decrease in diversity in the fall.
Prior to restoration, guild structure was relatively constant across all seasons, with generalists most abundant in spring and summer and decreasing in fall (Figure 7). There were more birds in each of the six guilds after restoration, with a marked increase in generalist abundance in spring and summer and the addition of mudflat and open-water foragers across all seasons. Post-restoration spring and summer guild structure was somewhat similar, whereas fall abundances were heavily dominated by mudflat foragers. Abundance of open-water foragers remained somewhat similar across the three seasons, whereas generalists declined in the fall.
Post-restoration Avian Trends at Harrier Meadow and Mill Creek
Shannon diversity of habitat types at Harrier Meadow (1.12) was only slightly higher than at Mill Creek (1.09), despite the considerable difference in habitat configuration (Figures 2 and 3; Tables 1a and 1b).
Across the three years surveyed after restoration, cumulative species richness was 78 species (8 species per hectare) at Harrier Meadow and 65 species (4.6 species per hectare) at Mill Creek. Similarly, on a per-survey basis, Harrier Meadow also had a higher average number of species per hectare than Mill Creek in all years after restoration (Table 8). Within each individual marsh, species richness per hectare did not vary across post-restoration years. Density was also consistently higher at Harrier Meadow than at Mill Creek after restoration (Figure 8). Both marshes exhibited a decrease in avian density over time; however, the decrease was substantial only at Harrier Meadow, with density decreasing by 60% between 2001 and 2003. Though there was considerable variability, the largest consecutive interannual decline (59%) occurred between 2002 and 2003 at Mill Creek. At both marshes, the five species with the highest densities were Canada goose (Branta canadensis), mallard (Anas platyrhynchos), snowy egret (Egretta thula), great egret (Casmerodius albus), and gadwall (Anas strepera).
Click image to enlarge
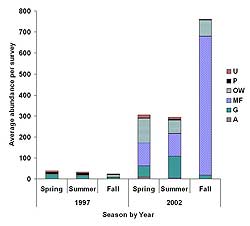
Figure 7. Average abundance per survey of six foraging guilds at Harrier Meadow before (1997) and after (2002) restoration during spring, summer, and fall. Guilds include upland foragers (U), Phragmites australis foragers (P), open-water foragers (OW), mudflat foragers (MF), generalists (G), and aerial foragers (A).
After restoration, avian diversity and evenness did not differ markedly between marshes in any year or among years at either marsh (Table 8).
Both Harrier Meadow and Mill Creek were dominated by three foraging guilds: open-water, mudflat, and generalist. While the relative proportions of guild density were very similar in 2001 at both marshes (Figure 9), the density of generalists, open-water, mudflat, and upland foragers at Harrier Meadow was more than double the density at Mill Creek (Figure 10). Average density of generalists decreased significantly between 2001 (10.6 ± 1.7) and 2003 (3.4 ± 0.7) at both Harrier Meadow and Mill Creek (2001: 4.6 ± 1.3; 2003: 0.89 ± 0.2). At Mill Creek, the density of open-water foragers significantly decreased between 2001 (6.2 ± 1.1) and 2003 (3.2 ± 0.7).
Migratory flocks of Calidris species sandpipers made a disproportionately large contribution to overall avian density at both Harrier Meadow and Mill Creek after restoration. In 2001 and 2002, sandpipers accounted for over 90% of mudflat-forager abundance and over 60% in 2003. Average flock size was similar at Harrier Meadow (488 ± 171) and Mill Creek (447 ± 184). The average flock size of sandpipers decreased at both marshes from 2001 (Harrier Meadow 874 ± 384; Mill Creek 809 ± 558) through 2003 (Harrier Meadow 191 ± 119; Mill Creek 108 ± 44).
Discussion
Several aspects of the study design proved to be of particular value in assessing the avian response to restoration at Harrier Meadow. Most notably, the pre-restoration data for Harrier Meadow allowed us to make a direct, same-site comparison of the avian community present in the pre- and post-restoration habitats. The exclusive use of baseline or pre-restoration avian data from a separate marsh might have introduced confounding factors such as landscape context and hydrology and complicated the assessment of how restoration affected the avian community. Studying avian community structure at multiple temporal scales (annual, seasonal, and multiyear trends) also proved to be informative because we were able to detect patterns in the avian community typically hidden at the annual scale. Furthermore, the use of foraging guilds allowed us to gain a better understanding of how changes in the availability of particular resources in the marsh may influence avian habitat use (Miller & Cale, 2000).
Click image to enlarge
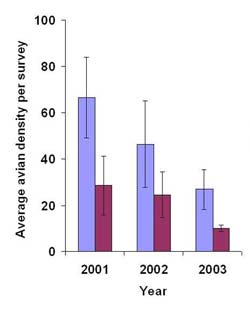
Figure 8. Average (± SE) avian density per survey at Harrier Meadow and Mill Creek during three consecutive years after restoration.
Pre-restoration Versus Post-restoration
Changes in avian community structure at Harrier Meadow reflected the change in habitat heterogeneity resulting from restoration. Prior to restoration at Harrier Meadow, breeding-season species richness was low (Burger, Shisler & Lesser, 1982), and abundance was dominated by a single species, the red-winged blackbird. This type of single-species-dominated community was consistent with avian communities found in other marshes with low habitat heterogeneity and a lack of surface water (Moller, 1975; Reinert, Golet & DeRagon, 1981; Craig & Beal, 1992).
Restoration of Harrier Meadow increased habitat heterogeneity considerably, including a large increase in open-water habitat. Avian community structure also changed considerably. The most prominent changes included a significant increase in avian abundance and an accompanying transition from a community dominated by passerines to one dominated by waterbirds. This type of avian response is supported by other studies that found a direct relation between the amount of surface water available in a marsh, avian abundance, and the use of the marsh by waterbirds such as waterfowl, shorebirds, and wading birds (Reinert et al., 1981; Burger et al., 1982; Slavin & Shisler, 1983).
The availability of open water at Harrier Meadow may have also affected the temporal dynamics of the avian community. After restoration, avian community composition was much less variable across consecutive surveys. The pre-restoration variability may have been caused by unpredictability of water availability in the marsh. With tidal influence restored and new ponds created, there was a greater area of surface water in the marsh, and the availability of water was more persistent throughout the year. As a result, avian species that relied on water for various activities, including foraging, were consistently present in the restored marsh, whereas they were occasional visitors in pre-restoration surveys.
Temporal Avian Responses to Restoration
Click image to enlarge
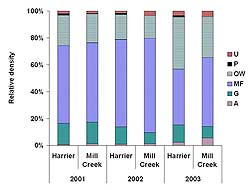
Figure 9. Relative density of six foraging guilds at Harrier Meadow and Mill Creek after restoration. Guilds include upland foragers (U), Phragmites australis foragers (P), open-water foragers (OW), mudflat foragers (MF), generalists (G), and aerial foragers (A).
The density of open-water foragers at Harrier Meadow was more than double that at Mill Creek, despite the fact that Mill Creek contained a greater area of open water. This implies that factors other than habitat availability per se influenced avian use of open water in the marshes.
This study did not include a means of rigorously determining what these factors were; however, one possible explanation became apparent during the study, and it has implications for future restoration design: Greater habitat heterogeneity and the configuration of the habitat mosaic at Harrier Meadow may have been responsible for the greater density of open-water foragers observed there. Phragmites and high-marsh grasses grew along the perimeter of open-water areas and extended outward at Harrier Meadow, creating protective cover and resting habitat for wading birds, shorebirds, and waterfowl (Reinert & Mello, 1995).
At Mill Creek, however, the band of vegetation along the perimeter of open-water areas was narrow compared with that at Harrier Meadow, and there were no large expanses of high marsh. Mill Creek therefore had limited resting area and protective cover for birds, possibly leading to limited use of open water. Other factors not addressed in this study, such as water depth, food availability, adjacent land use, and wildlife management may also have contributed to differential use of the two marshes studied.
In the mudflat foraging guild, the average flock size of Calidris species sandpipers decreased at both marshes from 2001 through 2003. During the same time period, but at Harrier only, we also saw a decrease in green-winged teal (Anas crecca), another species that forages on the mudflat in both marshes during migration. Concurrently, vegetation became established on what were previously bare mudflats, a phenomenon that has been linked to a decrease in avian use (Eertman, Kornman, Stikvoort & Verbeek, 2002). We do not believe the vegetated areas were large enough to cause the observed decrease in sandpiper abundance and suggest this as an interesting focus for future research. Several species in other guilds also decreased in abundance during the post-restoration monitoring period. In the open-water foraging guild, for example, mallard decreased at both marshes while great and snowy egrets decreased at Mill Creek.
Click image to enlarge
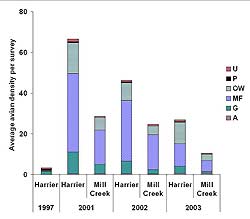
Figure 10. Average density per survey for six foraging guilds at Harrier Meadow and Mill Creek after restoration. Guilds include upland foragers (U), Phragmites australis foragers (P), open-water foragers (OW), mudflat foragers (MF), generalists (G), and aerial foragers (A).
It is possible that the observed changes in abundance at Harrier and Mill Creek were actually occurring on a scale larger than the local level and were not a result of post-restoration habitat changes in the marsh. For example, the decreasing trend in abundance of Calidris species sandpipers at Harrier Meadow and Mill Creek may not have been due to the loss of mudflat habitat at the restoration sites, but rather to population trends occurring at the broader, regional scale. Two sources of data would be useful in determining the scale at which changes in the avian community occurred: 1) regional-scale population data for the avian species observed at the restored marshes, and 2) avian community data from spatially independent, stable marshes within the Meadowlands. (We use the term "stable" to refer to a marsh that has not experienced a major disturbance, such as restoration or change in hydrologic pattern, for more than a decade.) If trends in the avian community at the restoration sites were similar to trends found outside the marsh, a "restoration effect" may be ruled out. Conversely, if trends within each restored marsh were unique to that locality, they may be interpreted to be a result of ecological dynamics associated with succession after restoration.
It appears that the restorations at Harrier and Mill Creek were successful because each newly created habitat was associated with increased avian use. However, with such a broad restoration goal as "to increase avian use of the marsh," it is very easy to deem both restorations successful. A positive or negative evaluation of these restorations would be more meaningful if it were based on more clearly defined goals. For example, the abundance of open-water foragers at Mill Creek increased considerably after restoration. This aspect of the restoration would be judged a success under the broad goal of increasing avian use of the marsh. If we take a closer look, however, we see that mallard accounted for 60% of the increase. If the goal were more specific in terms of target species, the general increase in open- water foragers might not be considered as successful. We stand to learn more about the restoration process by studying the success or failure to achieve clearly defined goals and realistic goals (Kentula, 2000; Choi, 2004).
In this study it was not possible to locate true reference sites for comparison with Harrier Meadow and Mill Creek. The urban context of the marshes placed numerous constraints on the restoration itself and on the evaluation of post-restoration progress (Grayson, Chapman & Underwood, 1999; Ehrenfeld, 2000). For example, the restoration plans had to take into consideration limitations due to habitat fragmentation and tidal restriction. Consequently, the marshes were not restored to the habitat composition typical of a salt marsh or any other naturally preexisting habitat type, hampering the use of reference sites. Moreover, the majority of marshes in the Meadowlands could not serve as reference sites as they are themselves ecologically degraded and dominated by Phragmites.
Few studies of avian response to marsh restoration in urban landscapes exist in the literature. This study serves as an essential first step toward developing a body of scientific research addressing avian response within the unique urban context. We suggest two particular areas in which future studies should be focused. First, our avian survey data showed differences between the two marshes in abundances of each foraging guild. Foraging studies directly linking birds and specific restored habitat types would provide insight into the mechanisms behind the success and failures in achieving restoration goals. Second, future studies should also investigate reproductive success in restored marshes in urban habitats. It is important to determine whether these urban marshes, which are under constant anthropogenic pressure (Ehrenfeld, 2000), serve as sources or sinks for the birds breeding on site.
While post-restoration monitoring has provided insights into the avian community response to marsh restoration, it is important to note that three years of post-restoration monitoring is a relatively short period from which to draw conclusions about the long-term trajectory of the marshes. As more long-term monitoring data become available, we are learning that it could take one to several decades for restored wetlands to recover ecological function, and that the time frame for a restoration is dependent upon the initial conditions of the wetland (Zedler & Callaway, 1999; Warren et al., 2002).
Acknowledgments
This study was supported by the New Jersey Meadowlands Commission and the New Jersey Agricultural Station. We would like to thank Robert DeCandido, one anonymous reviewer, and Rick Lathrop for their suggestions, which greatly improved the manuscript. We would also like to thank Niall Dunne for editorial assistance and Elaine Drazek and Andrew Krivenko for field assistance.