Patterns of Species Diversity in Eight Northeastern United States Cities
by Steven E. Clemants and Gerry Moore
Brooklyn Botanic Garden, 1000 Washington Ave., Brooklyn, NY 11225
Published online June 24, 2003
Abstract
In this paper, the native and nonnative floras of Boston, New York, Philadelphia, Washington, D.C., Detroit, Chicago, Minneapolis, and St. Louis urban areas are compared, and overall native diversity and nonnative diversity are correlated with a variety of factors. A total of 4,159 species has been reported in the eight urban areas. Of these, 2,708 (65.1%) are native to one or more of the urban areas and 1,451 (34.9%) are nonnative. Only 316 (11.6%) of the native species and only 109 (7.5%) of the nonnative species are common to all of the urban areas. When the similarity of native species is compared, Boston, New York, Philadelphia, and Washington, D.C., form a cluster, as do Detroit, Chicago, and Minneapolis; St. Louis is least similar to the other seven urban areas. Correlating climatic variables (growing season, temperature) and geographical variables (area, latitude, longitude) with species richness showed that nonnative species richness was most strongly correlated with longitude (probably as a function of age of settlement). This is in contrast with past research on native species showing a strong correlation of native species richness with latitude and elevations due to climatic differences present at different latitudes and elevations. Further studies that incorporate data from additional urban areas are needed to determine if nonnative species richness continues to be strongly correlated with time of a city's settlement.
Introduction
Patterns of species richness have long interested biologists (Wallace, 1878). The persistent and predictable patterns, even though actual numbers of species in the studies may vary, suggest that there are underlying, controlling factors. What those factors are and how they affect species richness has been the subject of numerous papers (Barthlott & Mutke, 2001; Currie, 1991; Currie & Paquin, 1987; O'Brien, 1998).
The best-known and most studied pattern is the latitudinal gradient. It has been demonstrated that there is a strong negative correlation between latitude and species richness for a number of species (i.e., as one samples in higher and higher latitudes, species richness diminishes [Fischer, 1960]). Other patterns that have been recognized are strong correlations between species richness and longitude, precipitation, temperature, potential evapotranspiration (PET), and insolation (sunlight) (Currie, 1991; Currie & Paquin, 1987; Palmer, 1995; Barthlott & Mutke, 2001). In North America, distinct latitudinal and longitudinal patterns have been recognized for several groups of organisms, including trees (Currie & Paquin, 1987), mammals and amphibians (Currie, 1991), and vascular plants (Palmer, 1995). These patterns can be modeled using climate data (O'Brien, 1998).
Urban areas are known to have different climates, soils, hydrology, etc., than nonurban areas (Pickett et al., 2001). One would therefore expect the patterns of species diversity to be different in these areas. Pysek (1998) compared 54 central European cities and concluded that the "occurrence of native and alien species in urban floras follows rather different pattern(s)."
One aspect of species richness that is particularly relevant in urban areas is the occurrence of nonnative species. In nonurban areas, Lonsdale (1999) found that 70% of nonnative species richness could be accounted for by three factors: native species richness, whether or not a site was a preserve, and whether a site was on an island or mainland. Similarly Stadler et al. (2000) found a correlation between nonnative species richness and native species richness. In urban areas, Pysek (1998) found that city size (area and population) was the best predictor of nonnative species richness.
This study was conducted to examine patterns of species richness in eight large northeastern United States cities and to determine what relationship there is between these floras and various factors that might be influencing native and nonnative species diversity.
Methods
Urban Areas
The urban areas in this study are defined as all the contiguous counties in and around the city with more than 86% of their populations living in urban areas, as measured by the United States Geological Survey (2000). The only exception is the New York urban area: We excluded Hartford and New Haven Counties in Connecticut and Hampden County in Massachusetts because including them would have greatly expanded the size of this urban area (already the largest) and because these counties are outside the study area covered by the New York Metropolitan Flora Project (Moore, 2002).
Database 1
- Database 1 PDF (350 KB)
- Database 1 Excel (635 KB)
A database containing all species in the eight urban areas and their native/nonnative status can be found in Database 1 at right. Table 1 provides information on the boundaries and data sources for each of the eight areas studied.
Predictors
Data for various predictors were calculated or compiled from a variety of sources (Table 2). Latitude, longitude, and elevation information was derived from airports within each urban area (Santos, 2002). The land area was calculated by totaling areas of counties taken from the United States Census Bureau (2001). Climate data (growing season, temperature, precipitation) were gathered from three sources on the Internet: WorldClimate (2002), the National Oceanographic and Atmospheric Administration (NOAA, 2000), and Koss et al (1988). For most data, airports were used as the standard location within the urban area; however, we used the nearest available reporting station to gather growing-season data (see Table 1). Settlement dates are from Encyclopaedia Britanica (1997). Current population data came from the U.S. Census Bureau (2001). Historical population data came from the University of Virginia Geospacial and Statistical Data Center (1998), except for that of the District of Columbia, which was obtained from Gilmore (1996).
Potential evapotranspiration (PET) was calculated using the Thornthwaite equation, E = 16C(10Tm/I)a, where E is monthly potential evapotranspiration in mm, C is the daylight coefficient, Tm is the average monthly temperature in Celsius, and a is an exponent derived from the heat index (I). I = Σ(Tm/5)1.51 and a = (67.5 x 10-8I3) = (77.1 x 10-6I2) + (.0179I) + (.492). Temperature data came from NOAA (2000). Daylight coefficient was derived by adding the median day length for each month and dividing by 12.
Statistics
To compare the similarity of the native and nonnative floras of the eight urban areas, we calculated a Jaccard index of similarity (Ludwig & Reynolds, 1988) for the native and nonnative floras of each urban area. This index was then used to generate a cluster analysis of community similarity using the UPGMA (Unweighted Pair Group Method With Arithmetic Mean) program of the NTSYSpc (Numerical Taxonomy System, version 2.01) statistical package (Rohlf, 1997). Because it has been suggested that factors influencing nonnative species richness and native species richness are different (Pysek, 1998), separate analyses were conducted on native and nonnative species.
To determine which factors were most correlated with native or nonnative species richness, a Pearson correlation matrix was calculated using the natural logarithm of the native/nonnative plant species ratio, area, latitude, longitude, growing season, mean January temperature, and mean annual rainfall. Principal component analysis was also calculated using the same set of variables. These statistical tests were performed in Systat 10.2 statistical software (Systat, 2000).
This set of variables was chosen from a larger set of variables (i.e., those reported in Table 2). From this larger set of variables, only one was chosen from variable pairs that were significantly correlated. For example, date of settlement was strongly correlated with longitude. Therefore, only longitude was used on the correlation analysis reported in Table 3.
Results
Floristic Similarity
A total of 4,159 species have been reported as occurring in the eight urban areas. Of this total, 1,451 (34.9%) species are not native to any of the urban areas, and 2,708 (65.1%) species are considered to be native to one or more of the urban areas. The highest percentage of nonnative species is in Boston (45.71%), the lowest in Minneapolis (19.27%). See Table 2 for further details for each urban area.
although all urban areas are part of the Eastern Deciduous Forest Formation (Braun, 1950), they have relatively few species in common. A total of 316 (11.6%) native species are found in all eight urban areas, and 109 (7.5%) nonnative species are found in all eight urban areas; overall, 425 species (10%) are found in common.
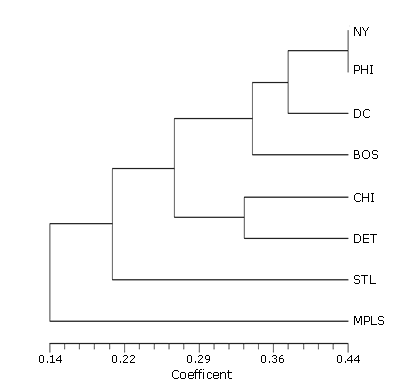
The cluster analysis of the nonnative species for each urban area (Figure 1) shows Minneapolis to have the most dissimilar flora, followed by St. Louis. The other two main clusters are Chicago-Detroit and Boston-New York-Philadelphia-Washington. In this second main cluster, Boston is the most dissimilar, followed by Washington (Figure 1).
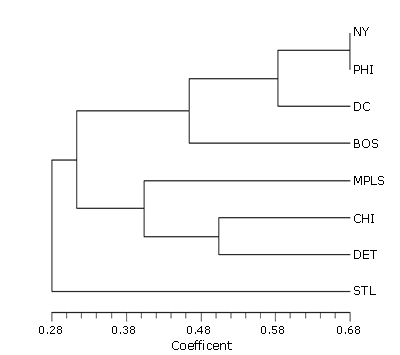
The cluster of the native species for each urban area (Figure 2) has St. Louis as the most dissimilar, followed by a Minneapolis-Chicago-Detroit cluster, with Detroit and Chicago again clustered together. The clustering of Boston, New York, Philadelphia, and Washington is the same as that in the analysis for nonnative species.
Factors Affecting the Native and Nonnative Flora Diversity
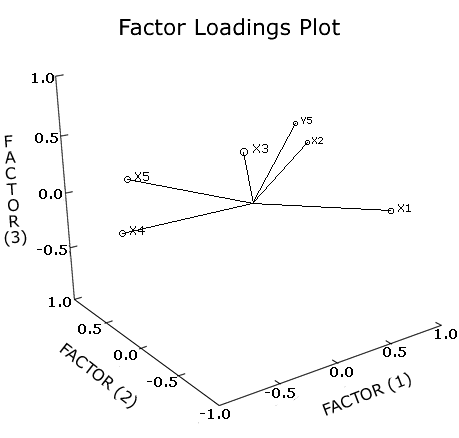
From the correlation matrix (Table 3) the natural logarithm of the native/nonnative species ratio was significantly correlated with longitude. Another significant correlation was between latitude and mean January temperature. In the factor analysis (Table 4, Figure 3), 96% of the variation is explained in the first three components (factors).
Discussion
Floristic Similarity
The similarity patterns of the native floras of the eight areas studied basically follow the current understanding of the vegetation of eastern North America. The first cluster (Figure 3), Boston, New York, Philadelphia, and Washington, are all part of the Oak-Chestnut Forest Region (Braun, 1950) and the Eastern Broadleaf Forest (Oceanic) Province (Bailey, 1995). These areas also have coastal-plain and shore floras not found in the inland cities. Detroit, Chicago, and Minneapolis are part of the Maple-Basswood Forest Region (Braun, 1950), whereas St. Louis, and to some extent Chicago, are part of the Oak-Hickory Forest Region (Braun, 1950). All four of these cities are part of the Eastern Broadleaf Forest (Continental) Province (Bailey, 1995).
While the cluster analyses based on native and nonnative species richness show some similar trends, there is one point in which they differ. Minneapolis is very similar to Chicago and Detroit in native species, but it has the least similarity of any of the urban areas when nonnative species are considered. This probably reflects the fact that Minneapolis is the farthest from a major seaport, where most nonnative species have been introduced.
Factors Affecting the Native and Nonnative Flora Diversity
The most striking correlation found in this study was between nonnative species richness and longitude. Currie and Paquin (1987) found a correlation between native tree diversity and longitude in North America, but no other study has found such a correlation. Though they don't present correlation statistics, Withers et al. (2000) show a defined longitudinal gradient for native and nonnative plant species in eastern North America. We believe that the nonnative species richness–longitude correlation found in our study is most likely the result of the history of plant introduction into North America. Longitude is strongly correlated with date of settlement. Furthermore, the cities that were settled earliest also are the cities with active seaports (all of them are on the East Coast). Most species were probably introduced via ocean ports along the East Coast and then spread inland. As a result, it is reasonable to conclude that seaports have the greatest number of nonnative species (as shown here) and that the number gradually declines as one moves inland.
Therefore, while we concur with the finding that factors influencing native and nonnative plant diversity are not the same, we have found different factors influencing nonnative diversity in North America than those found by Pysek in Europe. However, comparing our results with Pysek's (1998) is somewhat problematic because in his study—as in most Old World studies—species diversity was divided into native species (those that evolved in the region or arrived there before the Neolithic) and nonnative species (those that have been introduced to the region since the Neolithic); the nonnative species were further subdivided into archaeophytes (species introduced before 1500) and neophytes (species introduced after 1500). North American cities cited in this study have no archaeophytes because none were settled before 1500.
In Table 5, the results of this study are compared with Pysek's figures for 54 urban areas in central Europe (Pysek, 1998). It is evident that in our study the data samples for both population and area are much larger: Pysek included settlements of as few as 5,000 individuals, whereas our study included only settlements of more than 1 million individuals. The numbers of nonnative and native species in our study are much larger as well (although the percentage of nonnative species is in the same range as that for the European cities). From this we surmise that the floras of European cities are either depauperate compared with those of U.S. cities, or that area is a major factor in accounting for the more diverse floras. Pysek found a strong correlation between species diversity and area, which supports this latter supposition.
This history of plant introduction would also support the other strong correlations we found between nonnative species richness and the date a city was first settled (by way of the correlation with longitude). The longer a city has been in existence, the longer nonnative plants have had a chance to become established. In addition, the greater the shipping activity, the greater the numbers of plants coming into a port.
For neophyte diversity, Pysek (1998) found significant regressions with population, area, and population density (all multiplicative regressions), as well as with temperature (multiplicative regression) and altitude (exponential regression). In this study we found significant correlations between nonnative species diversity and longitude. Furthermore, elevation and settlement date were strongly correlated with longitude. We did not find significant correlations between nonnative species diversity and area, latitude, temperature, or growing season. In addition, potential evapotranspiration and population for the year 2000 were strongly correlated with latitude and area, respectively.
One possible explanation for these very different results is that we selected eight cities with populations of over 1 million, while Pysek selected 54 cities with populations ranging from 5,000 to 1.9 million. If there were a stronger population gradient among the U.S. cities, we might see the same correlations. But it could equally reflect a much longer time line for European urban development.
In summary, the results of this study lead to one significant conclusion: The factors influencing native and nonnative plant diversity in urban areas are not the same. Thus, the well-established correlations between climatic factors and native species diversity do not apply for nonnative species richness. Nonnative species diversity is more complicated and appears to be significantly influenced by factors regarding the settlement of the city (e.g., date of settlement, presence of a seaport).
The results of this study are preliminary and cannot be used to establish any general patterns regarding nonnative species diversity in North America. Future studies should include additional urban areas in North America to determine whether factors such as settlement date are causing the strong correlation noted in this study between nonnative species diversity and longitude.