Ruderalization in a Roman Park as a Result of Changing Management
by P.M. Bianco., G. Fanelli, P. Tescarollo, and S. Pignatti
Dip. Di Biologia Vegetale-Orto Botanico, Universitá degli Studi di Roma "La Sapienza," Largo Cristina di Suezia, 24 00165, Rome, Italy
Published online April 1, 2003
Abstract
Rome has one of the best open-space systems in Europe, and the Parco Regionale Urbano del Pineto is one of its crown jewels. The 250-hectare park is one of the city's last extensive undeveloped areas and has one of the most diverse floras in Rome. Presented here are the results of comparisons of studies conducted over a ten-year period of the vegetation in one-hectare quadrates situated in the park. We compared these studies in order to examine changes caused by a shift in land management. Once managed primarily as a sheep pasture, the park is now a nature reserve and used for recreation and tourism. This change has subjected the land to less pressure from animals and increasing pressure from humans. The comparison of the vegetation over time shows that natural succession has resumed, and reforestation is occurring in many areas. However, an analysis of Ellenberg's indicator values shows a clear increase in the number of ruderal species from the margins toward the center of the park. Neither trend bodes well for the future: Both reforestation and ruderalization will likely lead to a loss of biodiversity in the years ahead as important habitats and species niches are lost.
Introduction
Despite its rapid urban development, the city of Rome has one of most extensive green-space systems in Europe. Currently the park system, RomaNatura, includes 12 reserves, plus numerous historical villas, archaeological sites, and a number of gardens and uncultivated spaces (Cignini, Massari & Pignatti, 1995; Celesti Grapow & Pignatti, 1993).
Some of these parks are located in a buffer zone between the urban area and the surrounding countryside, forming a sort of greenbelt surrounding the urban area. Others are now completely inside the urbanized zone, and their use ranges from agricultural to recreational, as is the case of Parco del Pineto; these sites now represent a residual biotope, where patches of natural vegetation have somehow been saved from development (Celesti Grapow & Fanelli, 1991; Fanelli, 1995; Fanelli, Pignatti & Tescarollo, 2001; Pignatti, 1995).
Floristic lists are powerful tools in analyzing ecological patterns. Floristic composition can be analyzed qualitatively, but a quantitative analysis of floristic patterns is possible as well by means of Ellenberg's indicator values (Ellenberg, 1985; Pignatti et al., 2001; Thompson et al., 1993). A comparative study of urban biotopes by analysis of floristic patterns and historical data provides clues to understanding the effects of shifts in land management (Anzaldi, Celesti Grapow & Pignatti, 1992; Brandes, 1985; Catena, Macigno, Mulder & Pignatti, 1995; Celesti, Minichetti & Petrella, 1989; Kowarik, 1990; Kunick, 1982; Sukopp, Hejny & Kowarik, 1990; Horbert, Blume, Elvers & Sukopp, 1982). The Parco del Pineto is a representative case study because a large amount of historical and recent floristic information is available (Montelucci, 1953-54; Bianco, 1994) and because the biotope, although completely surrounded by intensively urbanized areas, preserves seminatural vegetation and a rich biodiversity.
Area of Study
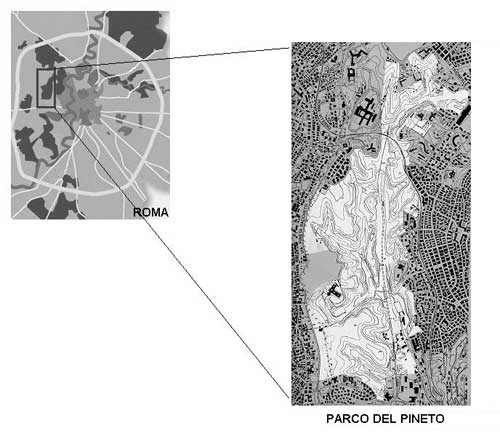
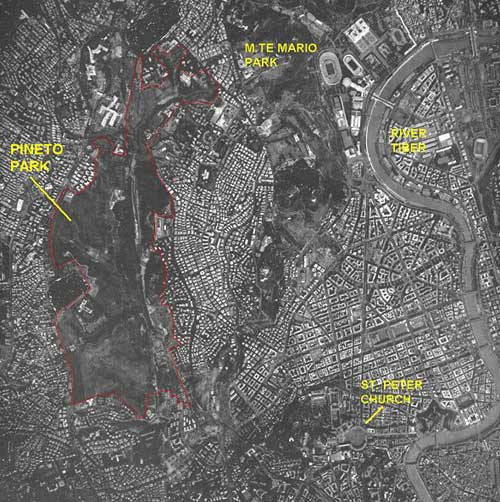
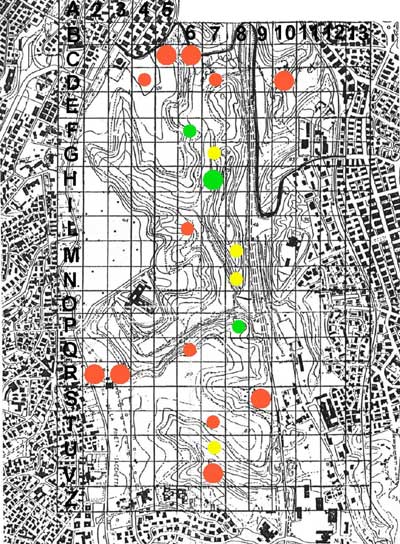
The Parco Regionale Urbano del Pineto is one of 15 protected areas within the city of Rome (Figure 1). The 247-hectare park is characterized by a remarkable variety of landscapes, including cork oak (Quercus suber) forests, species-rich grasslands, ponds, and wetlands; these diverse landscapes provide habitat for 642 plant species, giving the park the richest plant-species density in Rome (Celesti Grapow, Petrella, Fanelli & Lucchese, 1995) (Figure 2). The park is located within the metropolitan area, approximately two kilometers from the walls of the Vatican. Until about 15 years ago it was used primarily as a sheep pasture. Saved from development thanks to the efforts of citizens, it is the last expanse of undeveloped land in a highly urbanized sector of Rome. The park is in fact completely surroundend by heavily built-up areas with few green or open spaces (Figure 3).
Geomorphologically the Parco del Pineto is a tuffaceous plateau that rises above a lowland field (from which came the old place name "Vallis Infera"—Lower Valley). It is composed of small sandy hills (in part attributable to a system of fossil dunes) alternating with modest valleys. The geological substrata consist of four types or layers (Bonadonna, 1968; Carboni et al., 1991):
Mount Vatican (Unità di Monte Vaticano) (Vatican marls formation): clay and sands of the Pliocene-lower Pleistocene (3-1.8 million years ago). The clay increases toward the bottom of this layer and causes water accumulation in the valley bottoms;
Mount Mario (Unità di Monte Mario): silt and clay deposited during the lower Pleistocene (1.7-1.4 million years ago);
Mount Ciocci (Unità di Monte Ciocci): fluvial deltaic deposits of gravel and sand a little later than the Mount Mario substrate (1.3-1.2 million years ago);
Tufi Sabatini: deposits of tufa from the upper-middle Pleistocene.
In the valley bottoms, recent alluvial sediments are found.
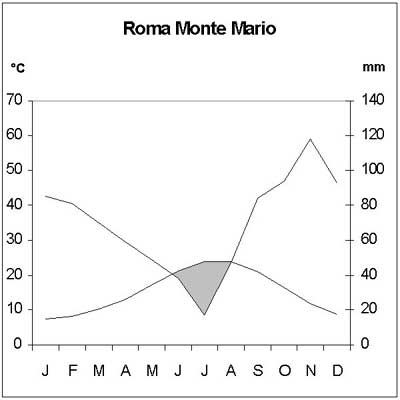
The area has a moderate Mediterranean climate, with a mean annual temperature of around 16°C and a mean rainfall of more than 800 millimeters annually. The average January temperature is 7.4°C, the average July temperature is 23.9°C, and frosts are very rare. There is a drought period that extends from June to August (Monte Mario meteorological station) (Figure 4).
The climate, the variety of substrates, and millennia of human activities have resulted in the area's great floristic diversity, making it an object of study since the last century (Montelucci, 1953-54; De Lillis & Testi, 1984, 1989; De Lillis, Testi, Scalfati & Cavendon, 1986; Bianco, 1994). Many rare species are present, in particular species from acidic soils poor in nutrients like Romulea rolli and Crocus suaveolens.
The vegetation of the area is very diverse, with about 15 vegetation types (Bianco, 1994; Fanelli, 2002). The primeval vegetation was probably a woodland dominated by the deciduous turkey oak (Quercus cerris) and Italian oak "Quercus frainetto" mixed with the evergreen cork oak "Quercus suber," a rare vegetation type that can still be found in relatively undisturbed sites in a few areas surrounding Rome. Wetlands were probably also present.
Nowadays the vegetation is dominated by Mediterranean species that have spread mainly as a consequence of burning and cutting. The main vegetation types include the following (Figure 5):
Quercus suber woodland: This is an evergreen woodland dominated by cork oak (Quercus suber) along with a few deciduous trees like downy oak (Quercus pubescens) and Italian oak "Quercus frainetto." The undergrowth is relatively rich with species like Stachys officinalis, Viola suavis, Smilax aspera, and the endemic Crocus suaveolens. This woodland is often subjected to fire, but due to its thick, corky bark, cork oak is very fire-resistant.
Corylus avellana woodland: This deciduous woodland is present as patches dominated by hazelnut (Corylus avellana). Other species are downy oak (Quercus pubescens) and poplar (Populus tremula). The undergrowth is scarce.
Dasypyrum villosum grassland: This grassland is dominated by tall annual herbs (1-1.5 meters) such as Dasypyrum villosum, Avena barbata, and Phalaris brachystachys. Perennials are also present, such as Asphodelus ramosus and Carlina corymbosa. This grassland is species-rich (about 30 species per ten square meters), including in particular annual legumes like Medicago polymorpha, Trifolium subterraneum, and Trifolium campestre. It has been irregularly subjected either to mowing (closer to the urban areas) or to grazing. Blackberry (Rubus ulmifolius), elm (Ulmus minor), and broom (Spartium junceum) thickets are sparsely present in the grassland.
Tuberaria guttata grassland: This is a low grassland (15-25 centimeters) growing on acidic soils, with a diverse floristic composition rich in rare or interesting species such as Tuberaria guttata, Crassula tillaea, Ornithopus pinnatus, Romulea rolli, and Rumex bucephalophorus. It is present in very small patches in a mosaic with the Dasypyrum villosum grassland and the Quercus suber woodland.
Wetlands: A few small springs in the park form ponds and ditches, where wetland vegetation occurs. The main species are Iris pseudacorus, Sparganium erectum, Lythrum salicaria, and Eupatorium cannabinum.
Perennial ruderal vegetation: This vegetation is present along the paths and on disturbed soils in moderate shade. It is dominated by nitrophilous forbs like Artemisia vulgaris, Cirsium arvense, Urtica dioica, and Melissa romana.
Annual ruderal vegetation: This vegetation is found as a narrow fringe along the paths that cut through the grassy areas. Main species are Hordeum murinum subsp. leporinum, Echium plantagineum, and Malva sylvestris.
Methodology
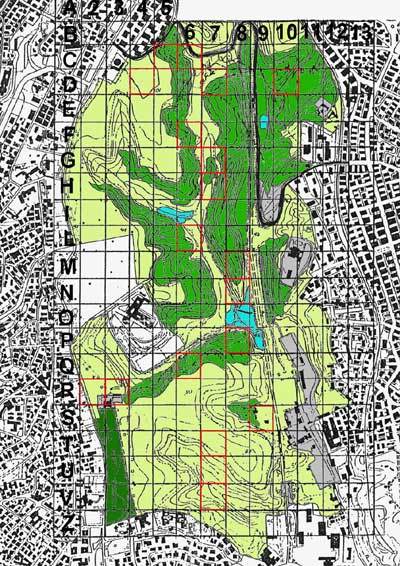
For the purposes of this study, the park was divided into quadrates of one hectare. Some of them, chosen randomly, were analyzed floristically in 1991 (Bianco, 1994) and again in 2000. In most squares a mosaic of different vegetation types is present. Quercus suber and Corylus avellana woodland is predominant in squares D7, D10, M8, N8, P8, and Q8. Dasypyrum villosum grassland predominates in the other squares, often together with small areas of ruderal annual vegetation, in particular in squares C6, C7, and R2. Wetlands are found only in squares P8 and N8. Nitrophilous vegetation is found in particular in squares Q6 and R3, but species typical of this type are found frequently in other quadrates, in particular C3, R2, and S8.
The quadrate floras (two surveys for each square, one in 1991 and one in 2000) were subjected to multivariate analysis, or centered Principal Component Analysis (PCA), using Sin-Tax 5.01 software (Podani, 1994) in order to identify variation. Moreover, each quadrate was analyzed according to Raunkiaer life-forms (1907, 1934) and Ellenberg's indicator values (1985). Differences between years have been tested by means of the paired samples T test. Raunkiaer life-forms have been calculated as percentages, and Ellenberg values as the average value of the species present in the square, without weighting abundance. Taxonomic nomenclature follows Pignatti (1982).
Results
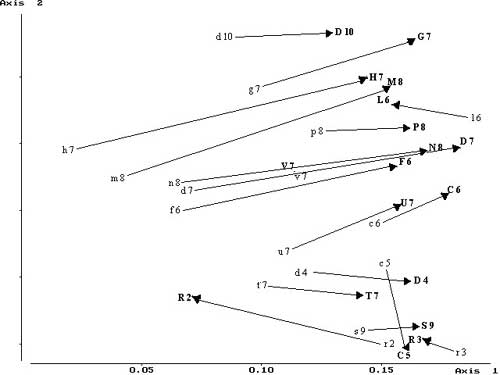
The main floristic trends are represented on the first two axes of the PCA diagram, which indicate a 67% variance. Along the first axis, surveys are shifted to the right from 1991 to 2000 (Figure 6). This shifting can be interpreted in light of the increase in perennial species and a decrease in annuals.
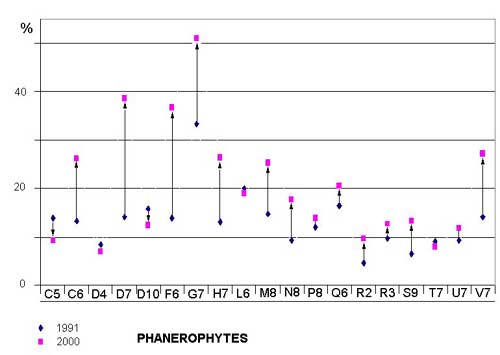
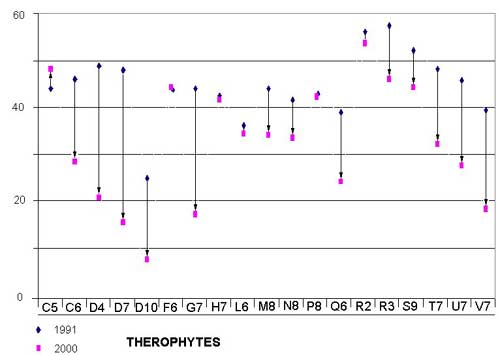
Phanerophytes (trees) and nanophanerophytes (trees and shrubs 0.25-2 meters), in particular Prunus spinosa, Ulmus minor, Rubus ulmifolius, and Crataegus monogyna, have increased to the detriment of the therophytes (annuals), which have diminished over time in nearly all the examined quadrates (Figure 7; Figure 8; Table 1). Differences between years are significant (t statistic -2.367, df 18, P = 0.029 (2-tailed).
Only quadrates C5, L6, R2, and R3 show a reverse trend. The surveys from 1991 are shifted to the left in 2000, and little variation or an increase in therophytes has been observed. These quadrates are near the main entrances to the park, where the growth of trees and shrubs is hindered by strong human pressure, such as trampling or cutting.
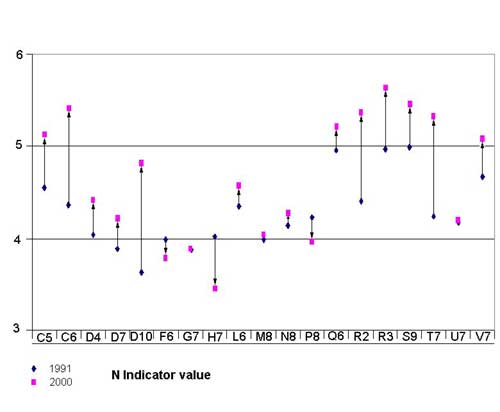
The second axis separates quadrates of the outer zones (C5, R2, R3, etc.) from those of the inner part of the park (G7, M8, P8, etc.). From 1991 to 2000 there was a slight shift downward of outer quadrates and upward of inner quadrates. This pattern on the ordination diagram matches Ellenberg's indicator values (Figure 9; Table 2), which fall into three distinct categories: quadrates in which an obvious increment was found (C5, C6, D10, R2, R3, S9, and V7), quadrates in which a moderate increment was found (D4, D7, L6, Q6, and T7), and quadrates in which the average of the indices remained the same (G7, M8, N8, and U7) or decreased (F6, H7, and P8). The distribution of the three categories indicates an increase of nitrogen indicator values toward the marginal areas of the park and a slight decrease in the inner zones. The overall change of nitrogen indicator value is highly significant (t statistic -2.367, df 18, P < 0.0001).
A closer study of the new species and those that disappeared suggests an explanation of the floristic changes. In quadrates where there is a strong increase in the nitrogen indicator values, species appear that are both nitrophilous and intolerant of competition, such as Arctium minus, Picris hieracioides, Sonchus tenerrimus, Urtica membranacea, and ephemerals such as Cardamine hirsuta and Fumaria officinalis. At the same time, oligotrophic annuals (those characteristic of areas low in nutrients like nitrogen) such as Aira cupaniana, Briza maxima, Petrorhagia prolifera, Trifolium arvense, Trifolium cherleri, and Trifolium echinatum disappeared. This is a clear indication that ruderalization is under way.
In areas with a moderate increase in nitrogen indicator values, there is an increase in only a few nitrogen-demanding species (Elymus repens, Arctium minus, Veronica polita). The main change associated with this indicator value is the disappearance of oligotrophic species such as Medicago truncatula, Plantago psyllium, Rumex bucephalophorus, and Tuberaria guttata.
Discussion
Since the first vegetation studies were done at Parco del Pineto, there has been a drastic change in management. In the past the park was mainly a sheep pasture, with moderate recreational use limited to the park's margins. The area was considered dangerous and "dirty" by the local people (Bonnes, De Rosa, Ardone & Bagnasco, 1989). In 1995, grazing was prohibited, and at the same time there was an increase in the number and frequency of visitors inside the park.
Other possible factors influencing the floristic composition of the Parco del Pineto include the use of certain areas for sewage works, railway tracks, and small public gardens, and the periodic burning of some areas. These are nonetheless limited to spotty areas scattered in the park and don't affect large segments of the landscape.
The most important result of these management changes is a general tendency toward reforestation throughout the park, as shown by the increase in phanaerophytes and nanophanaerophytes and the decrease in therophytes. The cessation of grazing has enabled vegetation succession to resume.
The data on the variation in nitrogen indicator values, as mentioned earlier, allowed us to distinguish three concentric areas in the park (Figure 10):
External Area: This includes areas near the border, which are subject to greater numbers of visitors and affected by construction. In this area, there has been a strong increase in nitrogen indicator values. This is associated with human impact, in particular the removal of shrubs, which favors the growth of nitrophilous forbs.
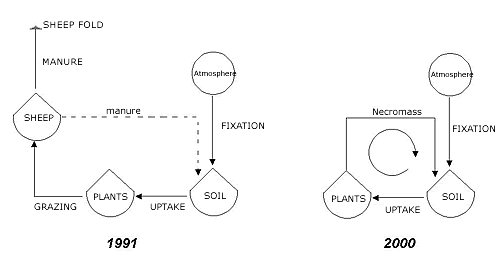
Intermediate Area: This area includes most of the lawns in the park and corresponds more or less with the zone that was grazed most intensively in the past. Here there has been a moderate increase in nitrogen indicator values, probably associated with the interruption of grazing. It is logical to conclude that with the disappearance of the animals there has been greater mobilization of nitrogen; nitrogen was once partially removed by the animals with grazing and only in part reintroduced in their excrement (Figure 11). All the nitrogen in the decaying vegetation is now available (Schmidt, 1978). The higher levels of nutrients have caused a gradual disappearance of oligotrophic species, which have been replaced by more competitive species.
Inner Area: This area includes most of the middle hill of the park and the less accessible areas; it also includes all those areas that were not heavily grazed. The nitrogen indicator values have remained the same, or in some cases, decreased.
The increase in nitrogen indicator values could also be affected by atmospheric deposits due to air pollution, a widespread phenomenon in recent years (Perakis & Hedin, 2002).
Conclusion
Our study demonstrates that the change in management of the Parco del Pineto from a pasture to a protected area for recreation and tourism has resulted in significant floristic changes. The abolition of grazing, which at first glance might be considered a good conservation measure, has coincided with an increase in nitrogen indicator values. This is possibly due to the fact that when grazing ceased, so did nitrogen removal by the sheep. Moreover, the increased number of human visitors has caused obvious ruderalization, especially in the more frequented outer areas of the park.
The future is not encouraging: We can expect a disappearence of oligotrophic, often rare species throughout the area and a decrease in biodiversity due to reforestation and the resulting reduction in the number of glades, microhabitats, and important species niches.
Acknowledgments
We greatly thank Dr. Andrea Palmeri and the members of the Cooperative Pineto 2000, Doctors Alessandro Serafini Sauli, Andrea Ramello, Diego D'Angeli, Giulia Rinieri, and Michele De Sanctis for their valuable contributions in the floristic surveys of 2000.